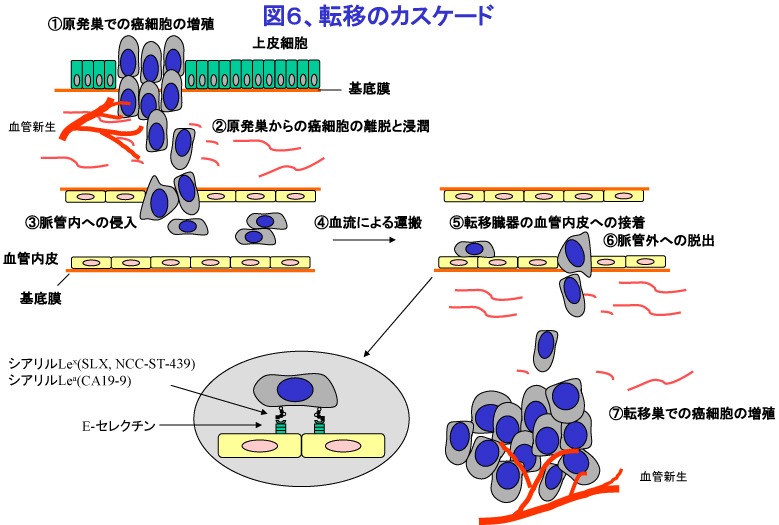
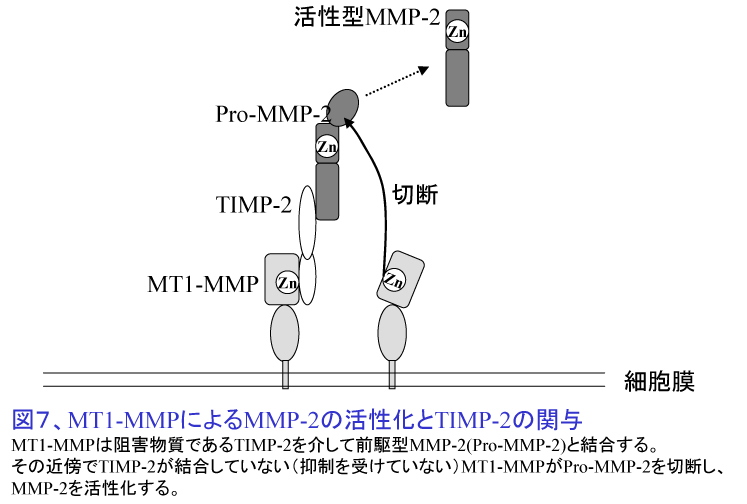
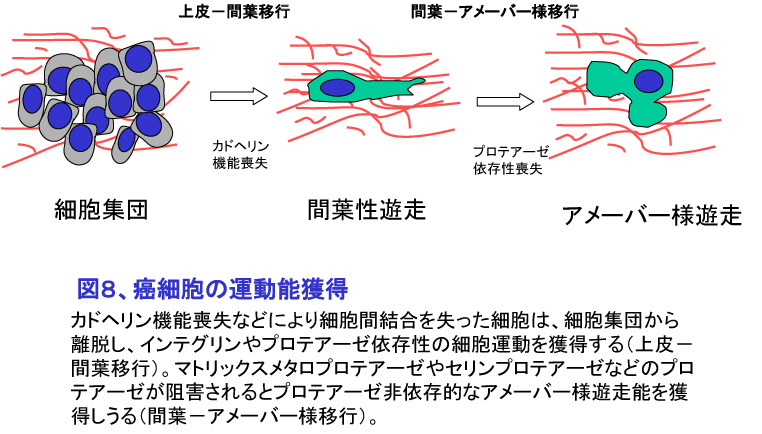
2. 転移の臓器特異性
癌種によって特徴的な転移臓器のパターンを示すことは臨床上よく経験するところであり転移の臓器特異性(organ tropism)と呼ばれている。転移の臓器特異性は、癌細胞を運ぶ血行動態と、癌細胞がある臓器で選択的に増殖しうる特性という2つの要素が関係している。臨床的には両方の要素が部分的に関与しあっている場合が多いと考えられ、相矛盾するものではない。前者は1928年にEwingより提唱された「転移の臓器特異性は血流の方向のみにより規定される」という考え方でanatomical-mechanical theoryとよばれている。消化器癌の肝転移はこの説がよく適合すると考えられる。後者は、1889年にPagetにより提唱されseed and soil theoryとよばれている。植物の種はいろいろな場所に植えられることができるが、それに適した土壌でのみ発育、成長することが可能であることから、癌細胞を種(seed)に、転移臓器を土壌(soil)になぞらえて、「癌の転移の成立は癌細胞の増殖に適した微小環境を有する臓器にのみ可能である」という考えである。近年、その分子機構が徐々に明らかになってきた。
1)ケモカインと臓器特異性
ケモカインは細胞に対して走化性(chemotaxis)を引き起こし、細胞間の相互反応を媒介するサイトカインで炎症反応を媒介する中心的因子である。ケモカインにはよく保存された4つのシステイン残基が存在し、N末端側に存在する2つのシステイン残基の間に存在するアミノ酸残基数により、CXC, CC, C, CX3Cケモカインの4つのサブファミリーに分類され、45種類以上が同定されている。ケモカイン受容体も20種類以上同定されている。
最近、(1)CXCR4を発現する癌細胞が、そのリガンドであるCXCL12を高発現するリンパ節、骨髄、肺に選択的に転移すること、(2)CCR7を発現する癌細胞が、そのリガンドであるCCL19, CCL21を高発現するリンパ節に転移すること、(3)癌間質中の線維芽細胞がCXCL12を産生して血管内皮細胞前駆細胞を癌組織中に呼び込むとともに、癌に対する増殖因子として働くことにより腫瘍進展に関与することが報告されたことにより、ケモカインやその受容体が転移の臓器特異性規定因子として注目され、それらの阻害薬による転移抑制効果が期待されている。
2)網羅的遺伝子解析による転移関連因子の探索
DNAマイクロアレイによる網羅的遺伝子解析やプロテオミクスによる網羅的蛋白解析が可能となり、2002年頃より癌の遠隔転移・再発に関係する遺伝子発現パターンが同
|