1. 原発巣での増殖
「増殖なくして転移なし」といわれるように、転移を形成するためにはまず癌細胞が原発巣で増殖する必要がある。原発巣での増殖には様々な増殖因子とその受容体がオートクラインまたはパラクライン的に関与していると考えられる。代表的な因子としてEGF(epidermal growth factor)受容体がある。EGF受容体は多くの固形癌に過剰発現されており、リガンド(TGF-a:transforming growth factor-aとEGFなど)が細胞外ドメインに結合すると、細胞内ドメインのチロシンキナーゼがリン酸化され細胞内シグナル伝達の結果DNA合成が刺激され、癌細胞の増殖を促進する。さらに、癌が1-2mm以上の大きさに増大するためには酸素や栄養を運ぶ血管の新生が必須であるが、血管新生も種々の血管新生促進因子と抑制因子により制御されている。血管新生については他稿で詳述されているので参照されたい。
2. 原発巣からの癌細胞の離脱と脈管への浸潤
増殖した癌細胞は次に原発巣から離脱する必要がある。これに関与する重要な分子としてE-カドヘリンが知られている。E-カドヘリンは細胞膜貫通蛋白であるが、細胞内裏打ち蛋白であるカテニンを介して細胞骨格に結合している。細胞外ドメインの先端で隣接する細胞のカドヘリンと結合することで細胞接着が生じる。すなわちE-カドヘリンは接着剤としての機能を果たしていて、その機能が失われると細胞同士の結合能力が失われる。高浸潤性の癌ではE-カドヘリンが低下し、原発巣から癌細胞が離脱しやすいと考えられている。
癌細胞が原発巣から離脱してもその周辺には細胞外マトリックス(ECM:extracellular matrix)が存在し、細胞の移動を妨げている。ECMは上皮下の間質結合組織とその境界面に膜状に存在する基底膜からなる。ECMは、各種コラーゲン、フィブロネクチン、ラミニンなどの糖蛋白質、プロテオグリカンなどを含んでいるが、主に基底膜はIV型コラーゲンで、間質はI型コラーゲンで構成されている。癌細胞は自身または間質細胞が産生する各種プロテアーゼやグリコシダーゼを利用してECMを分解し、生じた隙間を移動する。さらにECMへの接着、分解、移動を繰り返すことにより組織浸潤が達成される。これはECMとの接着にはインテグリンを中心とする接着分子が、ECMの分解には各種のプロテアーゼ(メタロプロテアーゼ、セリンプロテアーゼ、システインプロテアーゼ、アスパラギン酸プロテアーゼ)が関与しているが、その中でもメタロプロテアーゼ(MMP)が中心的役割を演じている。
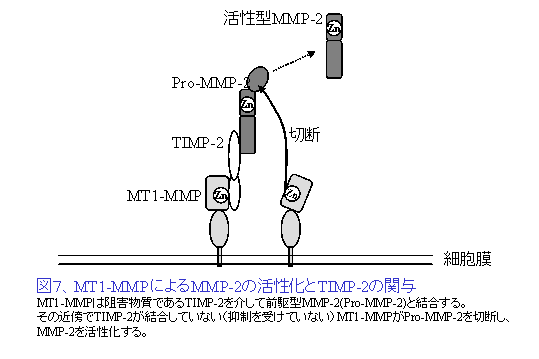
MMPは金属イオン要求性蛋白分解酵素で、25種類以上が同定されている。MMP存在様式の違いから分泌型と細胞膜型とに分類される。分泌型MMPはほとんどが細胞外に非活性型として分泌され、間質あるいは細胞表面プラスミンやトロンビンなどにより活性化され酵素作用を発揮する。よって、産生細胞から離れた広範囲のECMの分解が可能と考えられる。一方、膜型MMPは産生細胞の細胞膜表面に局在し、その近傍の基質を限定的に分解する。MMPの活性は、その発現量、活性化機構、あるいは生体におけるMMP抑制因子TIMP1-4との分子バランスなどによって厳密に制御されている。たとえば、膜型MMPのプロトタイプであるMT1-MMPは、代表的分泌型MMPであるMMP-2とTIMP-2をアダプターとして間接的に結合し、MMP-2を活性化する(図7)。これらの分子は癌細胞よりもむしろ線維芽細胞などの間質細胞が高発現し、癌の浸潤を促進していると考えられている。
![]()